ColabFold Tutorial
This tutorial introduces how to use ColabFold to predict protein structures with AlphaFold2, compare alternative multiple sequence alignment (MSA) strategies, and interpret model confidence carefully. You will work through monomer predictions first, then explore how custom MSAs and multimer predictions can change the results. By the end, you should be able to run ColabFold independently and judge when a prediction is biologically meaningful versus potentially misleading.
Exercise 1: Basic AlphaFold2 prediction
In this first exercise, you will submit a single protein sequence to ColabFold and generate an initial structure prediction. This gives you a reference model and lets you inspect the standard outputs produced by the notebook.
Target: PIGU subunit of the human GPIT protein (UniProt ID: Q9H490)
Input sequence: The FASTA file can be found in the Data section (Exercise 1), or you can copy it directly from UniProt.
-
Access ColabFold via GitHub - sokrypton/ColabFold and click AlphaFold2_mmseqs2.
-
Replace the default
query_sequencewith the PIGU sequence and rename thejobname(see Figure 1). - Run the next cell install_dependencies. It does not contain any modifiable parameters.
- Continue to MSA options. This cell controls the homology search for the input. Keep it at its default values and run the cell.
- The Advanced Settings cell allows for controlling the type of prediction model, recycling, MSA sampling and randomization. We will cover them later in this tutorial. For this exercise use its pre-loaded default setting.
- Run the Run prediction cell to start the prediction process (~17 min). The five computed protein models will be ranked by their predicted local distance difference test (pLDDT) score.
- Run Display 3D structure. Select the model to be displayed by its rank using
rank_num, then run the cell. This cell also allows for adjusting the colors and chain display. - Run the Plots cell to produce the predicted aligned error (PAE), sequence coverage and pLDDT plots.
- Save the results as a compressed zip file by running the Package and download results cell.
Tip
Alternatively, you can set all the parameters and then run all cells. Go to Runtime → Run all. Click "Run anyway" in the pop-up window to proceed.
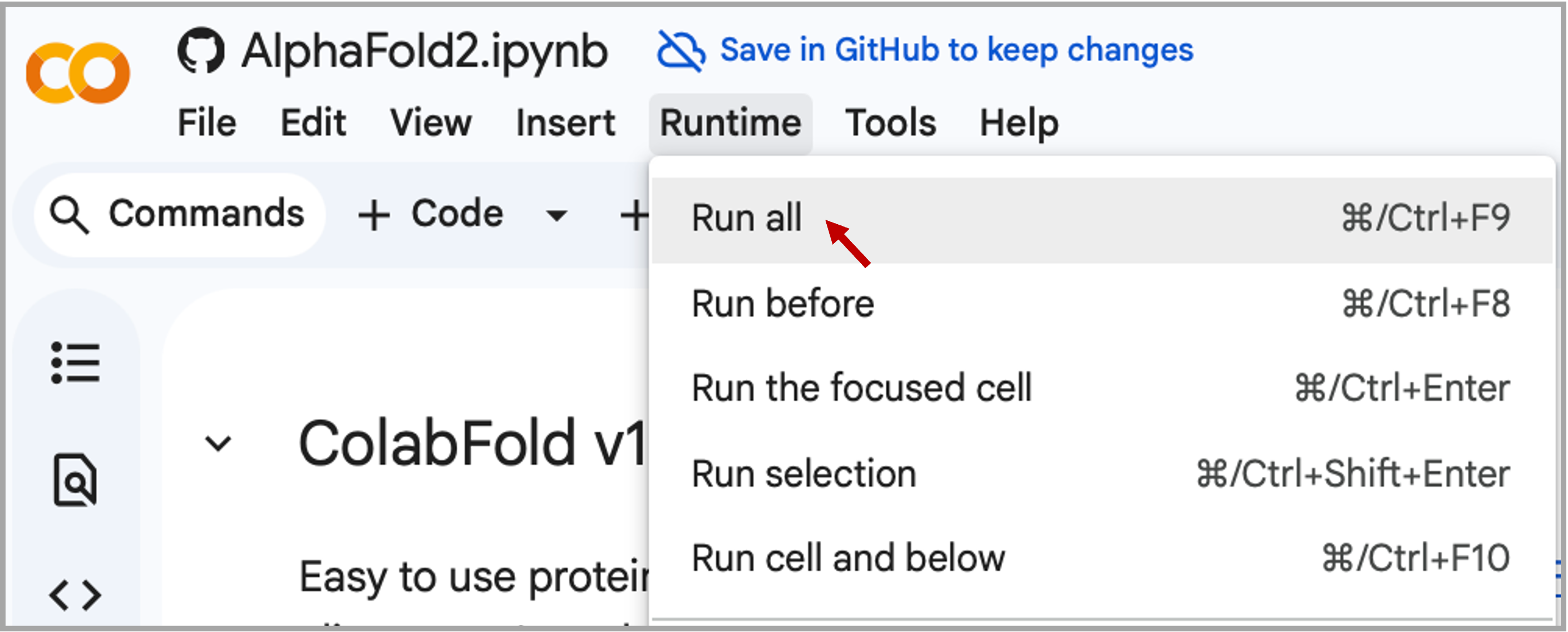
Warning
Google Colab typically allows only one active notebook at a time. Terminate any other Colab sessions before starting ColabFold. If you continue to have problems, or if Colab disconnects during the prediction, go to the "Runtime" menu, select "Disconnect and delete runtime," then click "Run all" to restart the process.
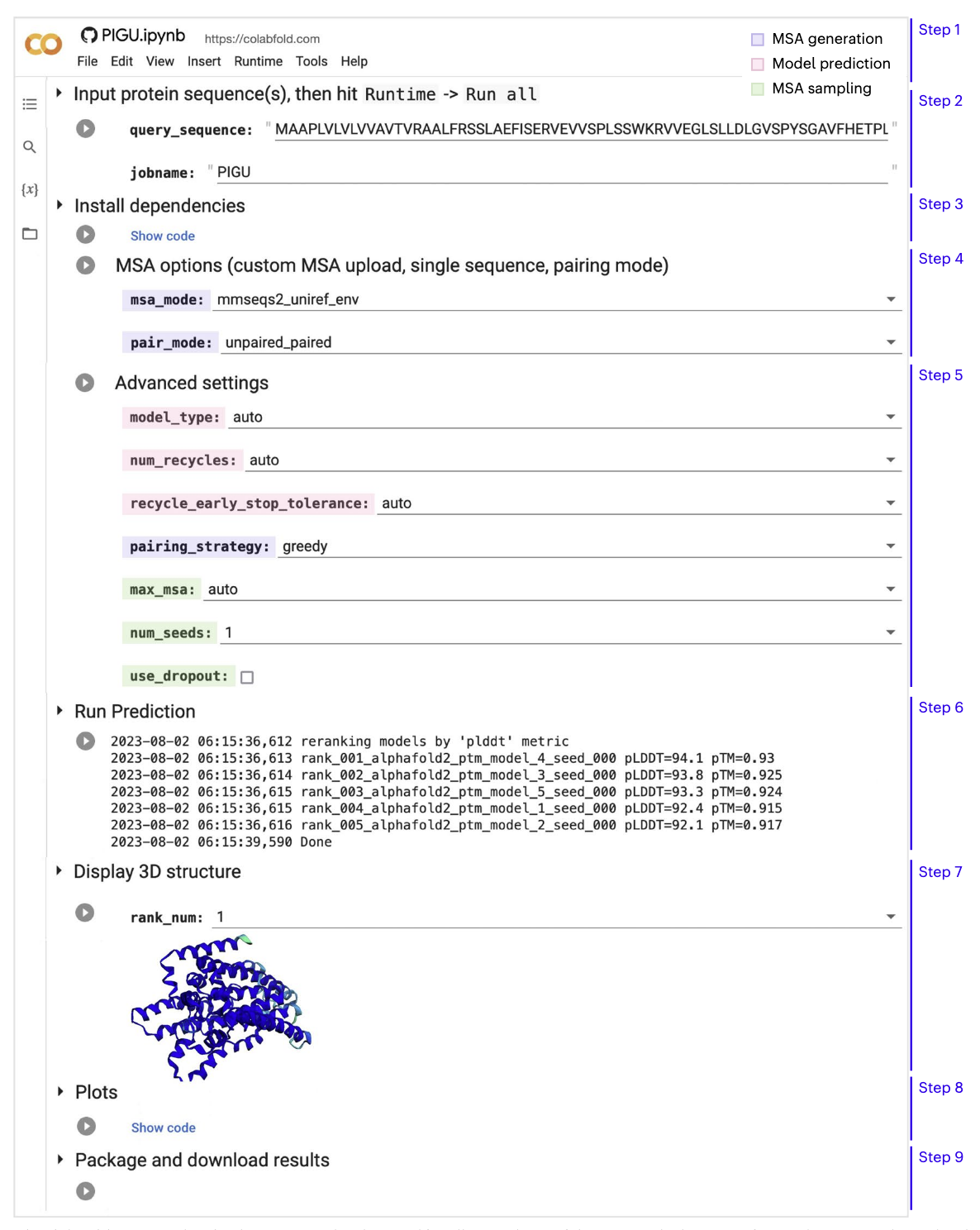
Web-based ColabFold-AF2 notebook. The figure from Kim et al., (2024).
Questions to consider
- Look at the sequence coverage plot. What can you say about the MSA quality?
- Examine and compare all five models. Is there agreement among them?
- Which regions are predicted with high confidence? Are there low-confidence loops or termini?
- Is there good coverage for these regions in the sequence coverage plot?
Exercise 2: Default versus custom MSA
In this exercise, you will compare the default ColabFold MSA against a custom MSA that you provide yourself. The goal is to understand how sequence depth and sequence diversity can affect structural confidence and model quality.
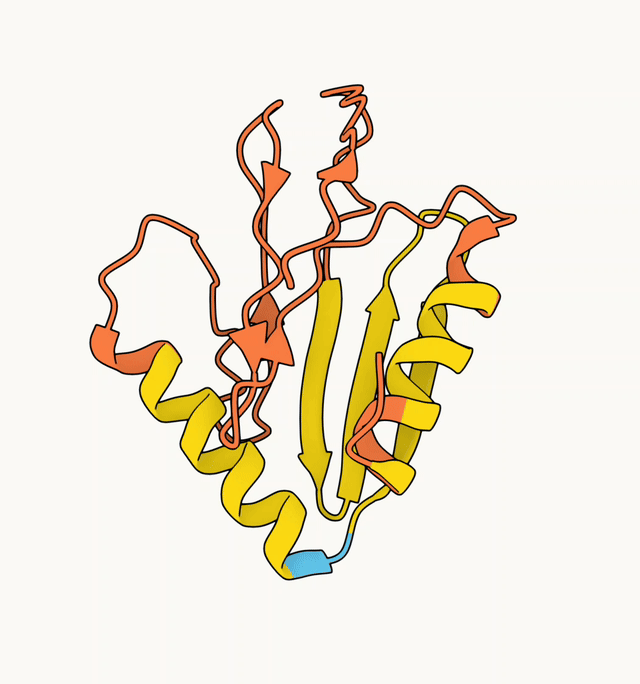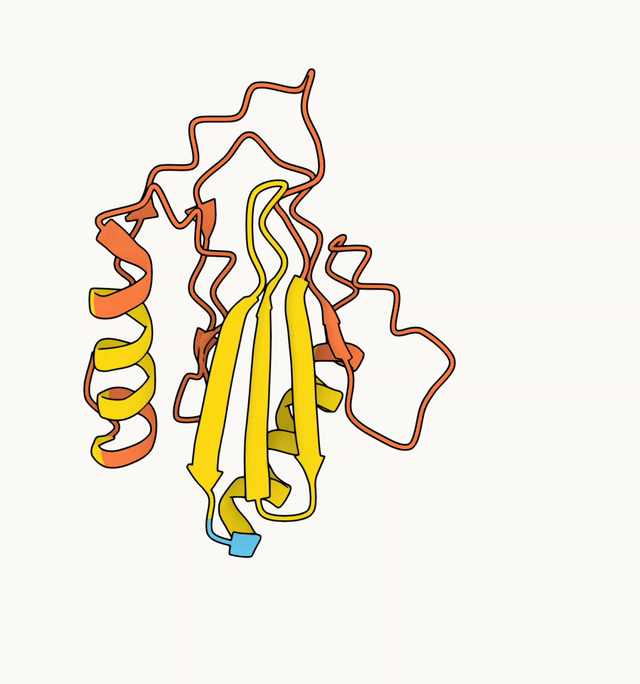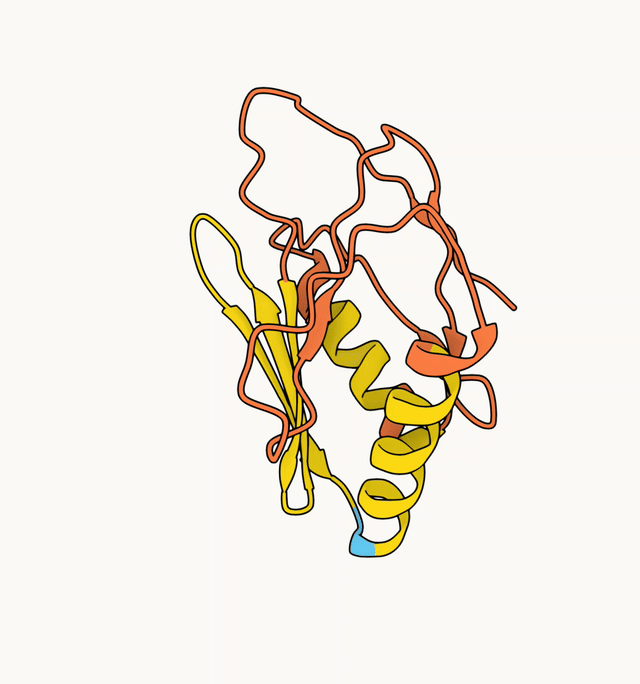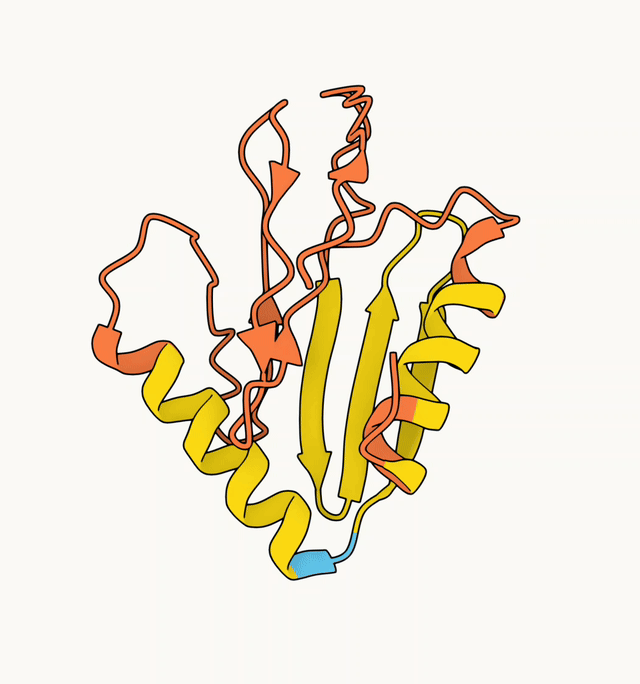

The AF2 model generated with the ColabFold default MSA is shown on the left (pLDDT 45.3), and the model generated with a custom HHblits MSA is shown on the right (pLDDT 89.2).
Target: Predicted viral protein (UniProt ID: G9B1X0)
Input sequence: The FASTA file can be found in the Data section (Exercise 2), or you can copy it directly from UniProt.
-
Run ColabFold with default parameters as described in Exercise 1. Evaluate the resulting models, confidence metrics, and plots showing sequence coverage and identity in the MSA. Save the results.
-
Prepare your own MSA in A3M format with HHblits Toolkit server:
Step 1: Paste the input sequence.
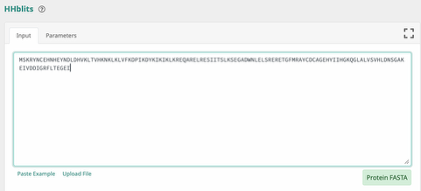
Step 2: Set the parameters as shown below and click Submit.
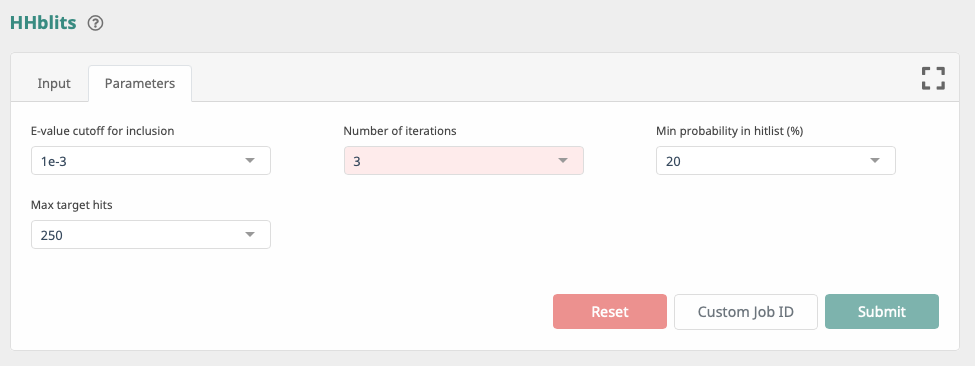
Step 3: Download the resulting MSA in A3M format.
Step 4: Open the downloaded file in a text editor and delete the first line containing the format marker
#A3M#, as shown below, otherwise ColabFold will return an error.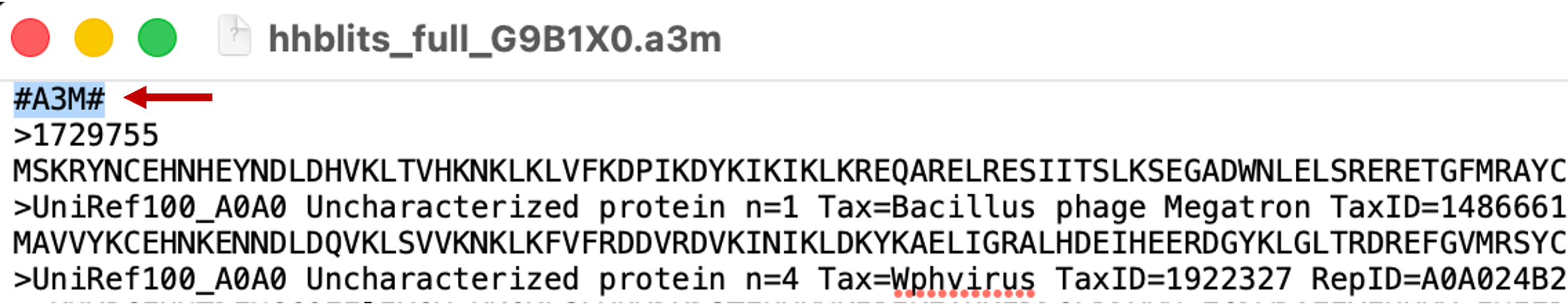
Troubleshooting
If your MSA source adds extra headers or nonstandard formatting, remove anything that may interfere with parsing before uploading.
-
Rerun ColabFold with the custom MSA: select
customformsa_mode, then upload the downloaded A3M file.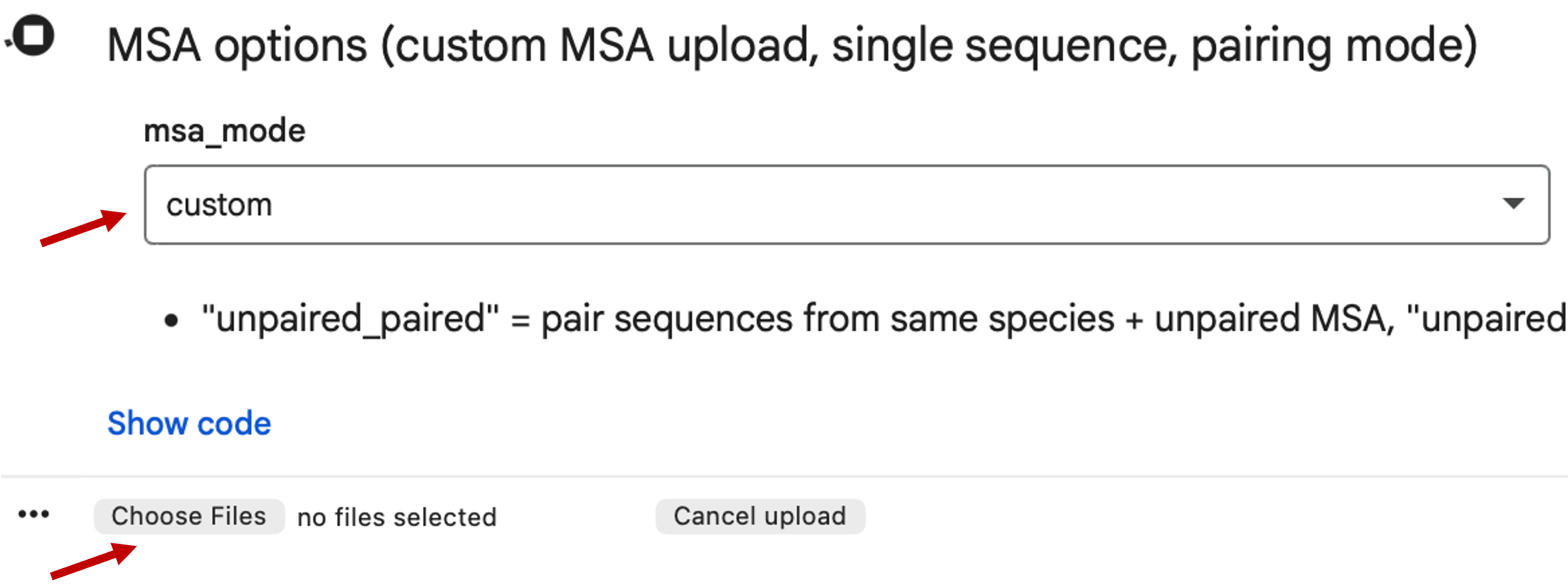
-
Evaluate the resulting models, confidence metrics, and plots with sequence coverage and identity in the MSA.
Questions to consider
- What is the difference between the ColabFold default MSA and the custom HHblits MSA used in this exercise?
- How do MSA depth, diversity, and coverage influence prediction confidence for this protein?
-
Save the results.
- Open the best-ranked models in Mol* and compare the structure generated with the ColabFold default MSA to the structure generated with the HHblits MSA. Superpose the models with TM-align and color them by pLDDT.
- Finally, check the AlphaFold Protein Structure Database (AFDB) entry for this protein and identify which MSA strategy produced the model most similar to the one shown in the AFDB entry.
Spoiler alert!
When comparing the sequence coverage plots for the default and custom MSAs, it is clear that the custom MSAs contain much more diverse sequences. This diversity was crucial for building a confident model of the target protein. The default MMseqs2-based MSA generation in ColabFold is fast, but it is sometimes not sensitive enough for difficult targets such as viral proteins. For highly evolved proteins or proteins with few homologs in the sequence database, and when the prediction with the default MSAs is not confident, it is recommended to generate a custom MSA for AlphaFold prediction using more sensitive sequence-search tools such as HHblits. For rare proteins, environmental sequence databases may also be more beneficial, as they can contain homologs that are absent from curated databases like UniRef, which are biased toward model organisms.
Exercise 3: Multimer prediction
In this exercise, you will examine a protein complex prediction. The aim is not only to run ColabFold multimer mode, but also to evaluate whether the predicted interface is likely to be real.
3.1 "Good example" for complex prediction
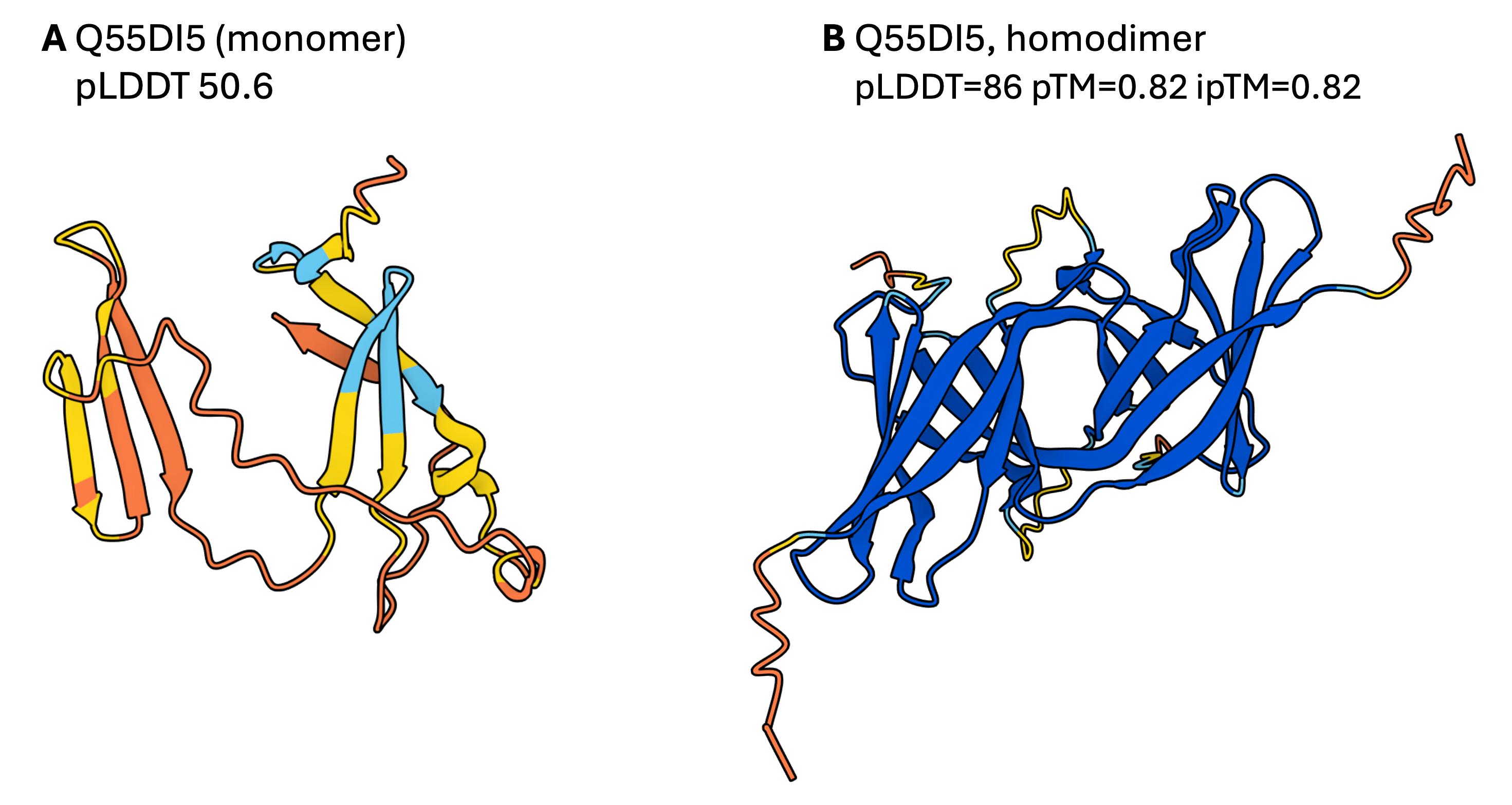
AF2 models generated with ColabFold in which the target protein was modelled as a monomer (A) and as a homodimer (B).
Target: Transcription elongation factor Eaf N-terminal domain-containing protein from Dictyostelium discoideum (UniProt ID: Q55DI5)
Input sequences: The FASTA file can be found in the Data section (Exercise 3.1).
-
Run ColabFold to model the target protein as a homodimer using the default parameters (~10 min). Use
:to specify inter-protein chainbreaks for modeling complexes. For examplePI...SK:PI...SKfor a homodimer.Note
You do not need to manually select AlphaFold-Multimer. The best complex prediction model (
alphafold2_multimer_v3) is selected automatically when theinput_sequencefield contains more than one sequence separated by:. -
While the complex prediction is running, open the AFDB and retrieve the monomer prediction for Q55DI5. Evaluate the model and its confidence scores. What can you say about them?
-
Download the mmCIF file for Q55DI5 from AFDB.

-
Once the ColabFold job finishes, evaluate the homodimer prediction and download the results.
-
Open Mol* and upload both the best ColabFold homodimer prediction and the AFDB monomer model. Superpose the monomer onto one chain of the complex. What do you observe?
Questions to consider
- What does the superposition tell you about the difference between the monomeric and dimeric folds?
- Why does the monomer appear disordered compared to the homodimer?
Spoiler alert!
As you can see for Q55DI5, the monomer has a lower average pLDDT and longer disordered regions, but the homodimer prediction reveals that the two chains intertwine, each contributing half a domain to form a stable fold. Therefore, to make a reliable prediction the domain requires the partner chain to fold correctly.
Questions to consider
But what is the native fold for Q55DI5? And what would you do if you don't know in which oligomeric state you should model the target protein?
-
Go to SWISS-MODEL Repository and paste
Q55DI5in the search field.
-
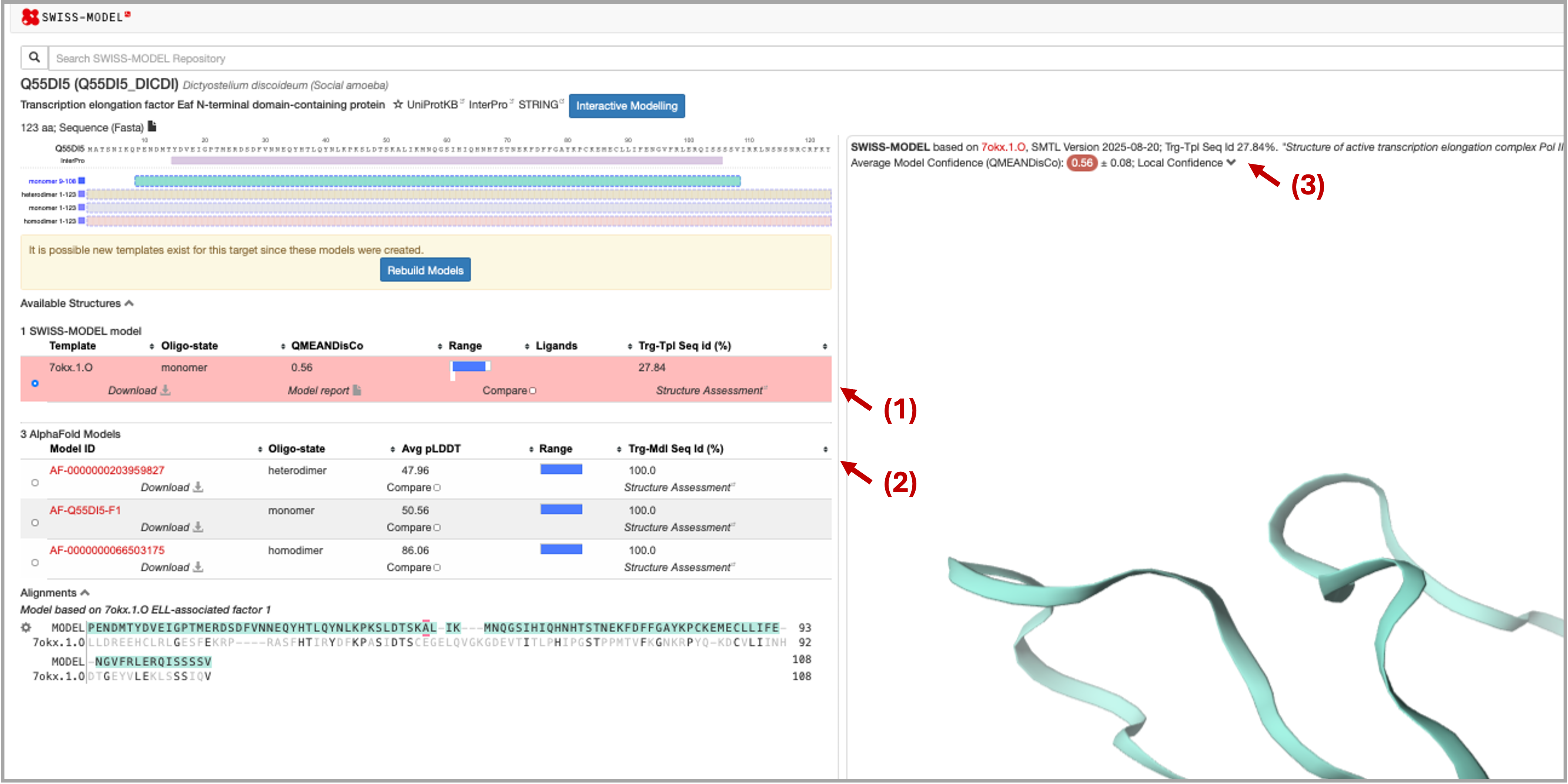
The results of the search show: (1) the best template found in PDB100 through a sequence search, which was used to build the SWISS-MODEL model (3); SWISS-MODEL also shows good hits found in AFDB. Chain O was the template from the found complex PDB ID: 7OKX (4).
-
Now superpose in Mol* the found template chain with the modeled homodimer. What can you say about that?
Spoiler alert!
Based on the found model, the target protein probably exists in a complex with other proteins, and the AF2 model for the homodimer is closer to the native fold than the monomer model.
3.2 "Bad example" for complex prediction
This exercise emphasises that AF2, like AF3, does not know the stoichiometry of the protein it models. Therefore, if we provide only a single sequence as input, AF2 will model it as a monomer (as seen in Exercise 3.1). The example below shows that a predicted monomeric fold is not always functionally meaningful, even when it has high confidence scores.
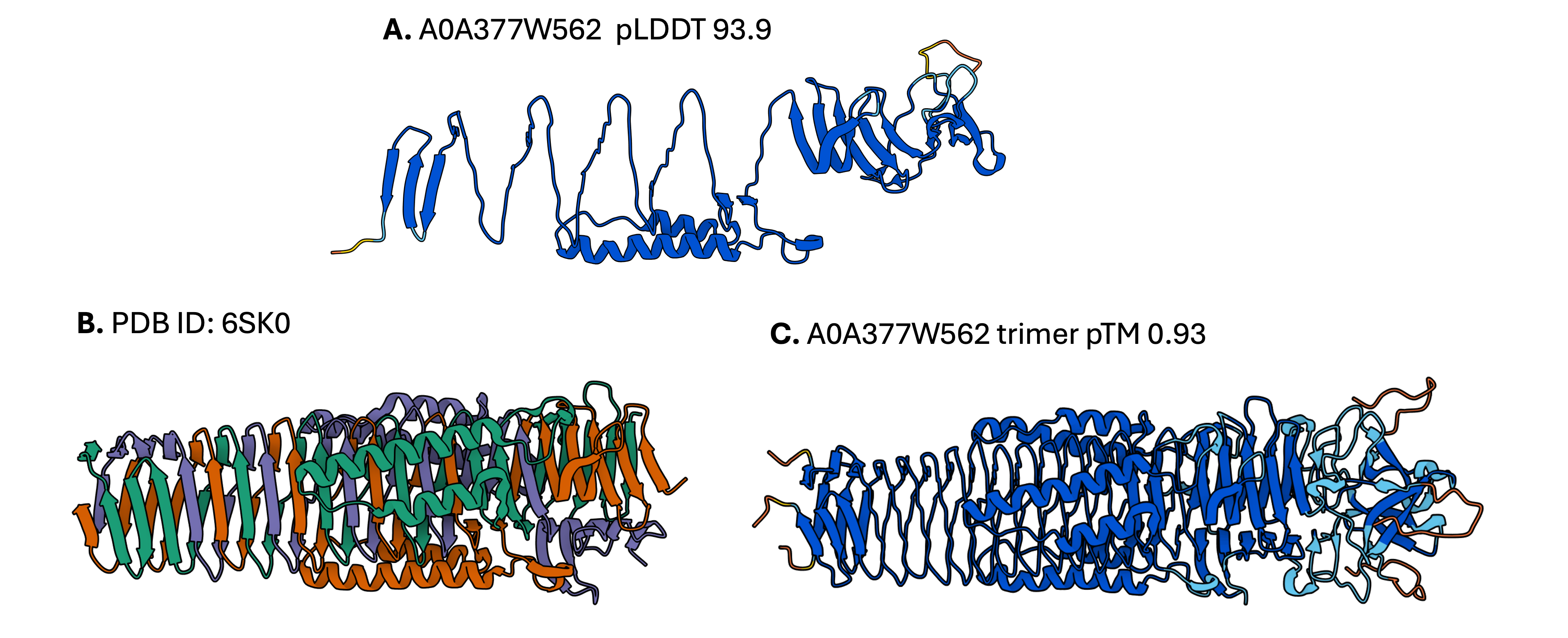
A. AF2 model of the target protein as a monomer; B. PDB structure of a homologous obligate trimeric protein from the E. coli Type VI secretion system; C. AF2 model of the target protein as a homotrimer. The figure is adapted from Durairaj et al. (2023).
Target: Type IV secretion protein Rhs from K. pneumoniae (UniProt ID: A0A377W562)
Input sequence: The FASTA file can be found in the Data section (Exercise 3.2).
- Run ColabFold with default parameters and model the target protein as a monomer (~13 minutes). To save time, the results are also available in the Data section.
- Evaluate the resulting models and plots. What can you say about the depth, diversity, and query coverage of the MSA? Look at the pLDDT and PAE plots — are they consistent across the 5 models? Overall, do you think the prediction is reliable?
- Now examine the predicted fold. Does it look like a native fold to you?
-
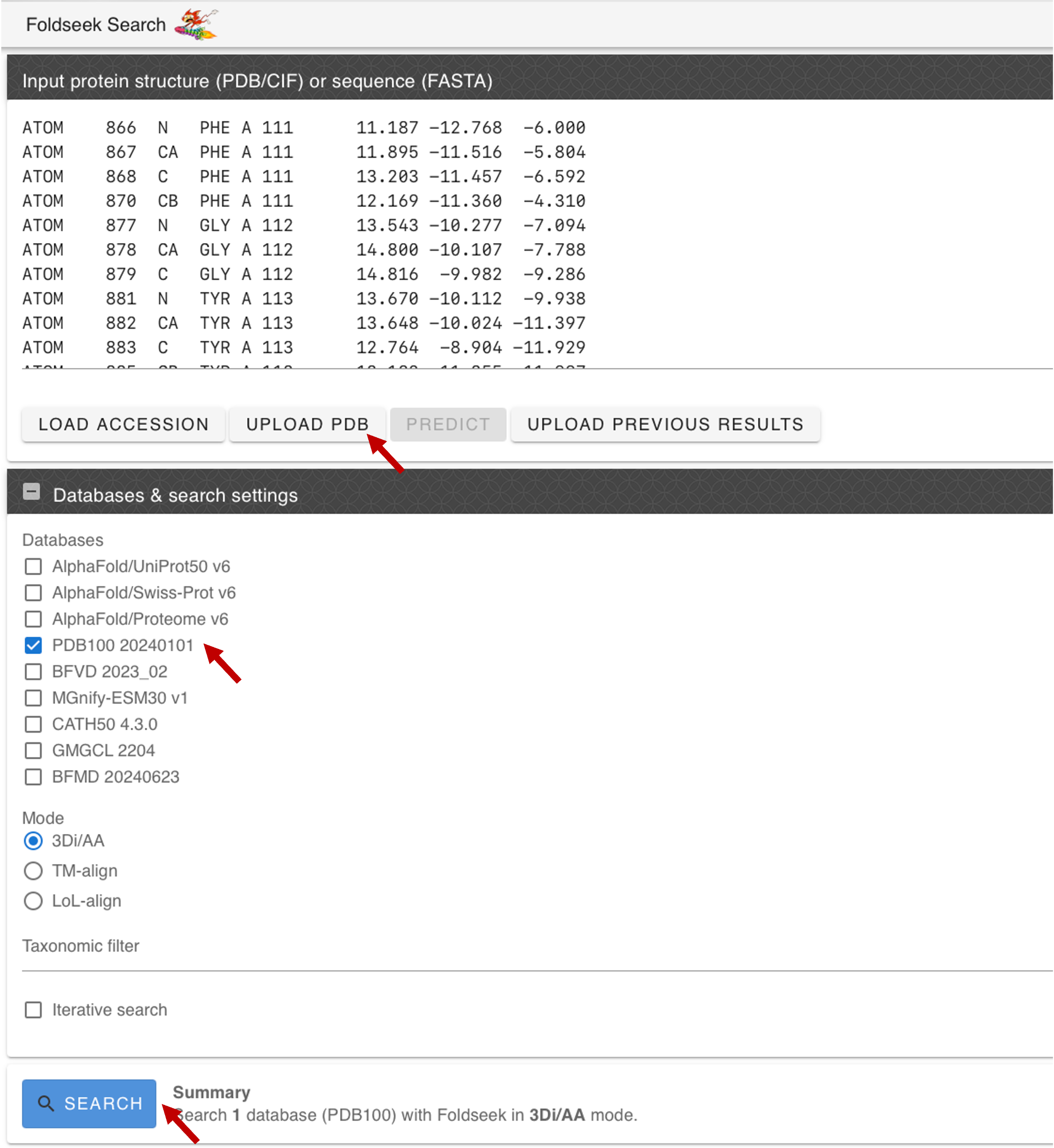
Go to the Foldseek server and search for similar folds in PDB100. Upload the PDB file of the best model, select PDB100 as the target database, and click Search.
-
Evaluate the results and click on the best hit: the VgrG spike from the Type VI secretion system. You will be forwarded to the PDB page with the experimentally resolved structure.
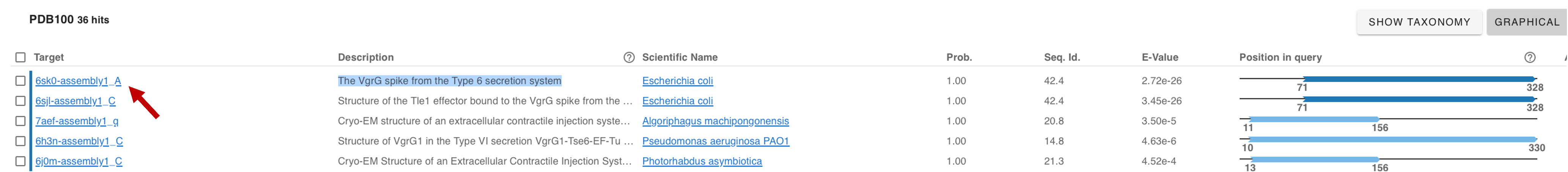
-
Open Mol* and upload the best monomeric model. Upload the Foldseek hit (PDB ID:
6SK0) structure using the Download Structure window in Mol*.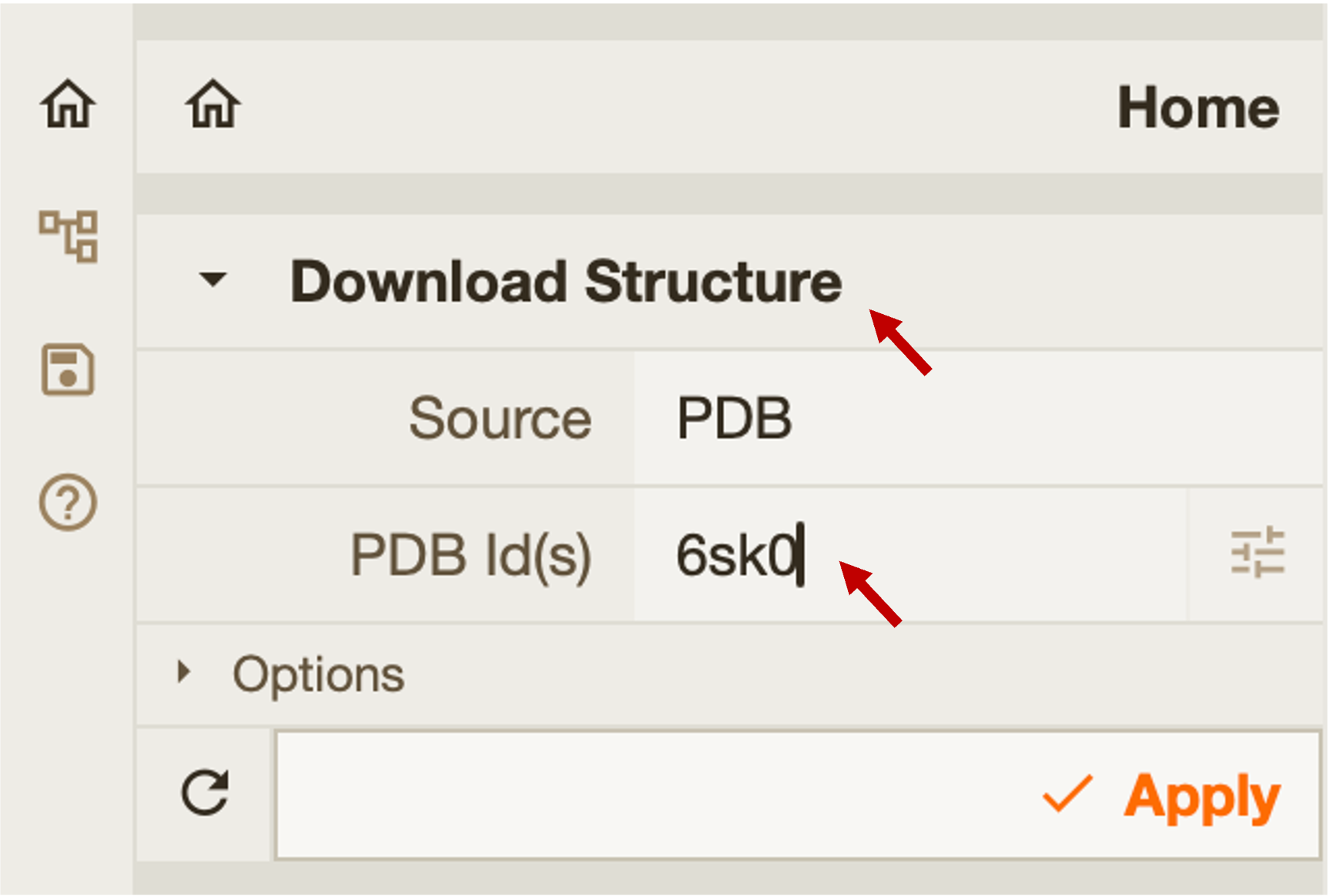
-
Superpose the AF2 model with the experimental structure. What do you observe?
- Consider whether the modelled protein might be part of a trimer rather than a standalone monomer. To test this hypothesis, paste 3 copies of the target sequence into the input sequence field, separated by
:, and run the prediction. This step takes approximately 1 hour; to save time, the results are also available in the Data section. - Evaluate the resulting plots and model. Superpose the predicted trimer with the experimental structure. Are there any notable differences in RMSD or TM-score?
Questions to consider
- Do the confidence scores alone tell you whether the monomeric fold is biologically meaningful?
- What conclusions can you draw from this exercise?
Spoiler alert!
AF2 does not model all conditions required for folding, such as the presence of binding partners. In this example, we used Foldseek to identify a homologous protein with a resolved experimental structure in order to infer the native oligomeric state of the target protein. The best hit identified by Foldseek is an obligate trimer, so the monomeric model obtained by ColabFold is unlikely to represent the native state, despite its very high confidence score. The AlphaFold-Multimer model of A0A377W562 as a trimer has very low r.m.s.d. to the PDB structure, which provides additional evidence for a trimeric oligomeric state of the target protein.
Ideally, this conclusion would be supported by low confidence scores for the monomer, as in Exercise 3.1, but that is not the case here. This may be explained by AF2 memorization, meaning that the model may have seen a related chain in a trimeric context during training, but we cannot say this for certain. See also the 4E-binding protein 2 example for a related case of AlphaFold memorization.
Exercise 4.0: Alternative conformations
During their normal function, transporters like OCT1 go through a series of conformational changes. This means OCT1 does not have a single canonical structure, but rather several. OCT1 has been solved experimentally by cryo-EM in two conformations:
The structure files are available in the Data section (Exercise 4.0).
- Open OCT1 on UniProt and copy the amino acid sequence.
- Run ColabFold with the sequence you just copied.
- Download the ground truth (experimental) structures in mmCIF or PDB format for both the inward-open state (8SC1) and outward-open state (8ET6).
- Compare the structure you get from AlphaFold to both of them (RMSD). To which one is it closer?
Exercise 4.1: MSA depth reduction or activating dropout layers
To sample different conformations (which is outside the initial scope of AlphaFold2) we have two strategies:
- Reducing MSA depth through the
max_msaparameter - Activating dropout layers by checking the
use_dropoutbox
In both cases the result also depends on the starting point (seed), so we can increase the number of times the model is run (by setting num_seeds to a bigger number), thereby increasing the chance some of them are in an alternative conformation.
Note
Running AlphaFold2 with more seeds (e.g. 16 instead of 1) produces more models (16×5 models instead of just 5), and takes much more time (16× longer). To save time, we will skip this exercise and try a different approach in the next exercise.
Exercise 4.2: Using templates
OCT3 is a different membrane transporter from the same family, which has recently been determined in its outward state (PDB ID: 7ZH0). We can provide this structure to try to bias the prediction of OCT1 towards the outward-open state.
The structure files (7ZH0 and 8ET6) are available in the Data section (Exercise 4.2).
- Download the outward state of OCT3 (7ZH0) in mmCIF or PDB format.
- In ColabFold, in the Input protein sequence section, change
template_modetocustom. When running the cell, it will ask for the template file (7ZH0). - The MSA AlphaFold constructs for this protein is deep and rich in data, so it will use the MSA heavily and ignore the template. That is why we need to reduce the depth of the MSA — in one of two ways (option a is recommended):
- Option a — Reducing the MSA depth: In the Advanced settings section, change
max_msato16:32or32:64. - Option b — Completely removing the MSA: In the MSA options section, change
msa_modetosingle_sequence.
- Option a — Reducing the MSA depth: In the Advanced settings section, change
- Compare the structures you get now to the ground truth structures (8SC1 and 8ET6). Structures should be more similar to 8ET6.
Spoiler alert!
In this exercise, we showed that AlphaFold2 can be guided toward alternative conformations, even though it was not originally designed for that purpose. One effective strategy is to reduce the MSA depth and provide a template representing the desired state. OCT3 outward-open template biased OCT1 toward an outward-open-like model, although the prediction remained somewhat intermediate. This demonstrates that homologous templates can help model alternative conformations, and tools such as SWISS-MODEL or Foldseek can be used to find suitable templates.
Exercise 5: Predict your favorite protein (bonus)
If you have completed all the exercises above, try applying ColabFold to a protein of your own choice.
Note
Prediction of long proteins or large protein complexes may take considerable time (~1–2 hours).
- Choose a protein and retrieve its sequence in FASTA format from UniProt.
- Check if there is a model available in AFDB; if not, run a first prediction using the default ColabFold parameters.
- Rerun the prediction using advanced options covered in the ColabFold lecture (slides available on the course website). Compare the two results: do the advanced settings improve pLDDT, PAE, or structural plausibility?
- (Optional) If you are not satisfied with the default MSA, build a custom MSA using the HHblits Toolkit server, download it in A3M format, and use it as input for a third run.
- (Optional) If you want to model different conformations, search for structural templates (e.g. using Foldseek or Swiss-Model) and rerun the prediction with custom templates and reduced MSA depth.